商机详情 -
深圳无需染色纺锤体胚胎发育
秋水仙素为什么会使有丝分裂的细胞停滞于中期
如果用秋水仙素处理有丝分裂的细胞,纺锤体会迅速消失,细胞停滞在有丝分裂中期,染色体无法分离成两组。用秋水仙碱进行诱导,从而将细胞阻断在细胞分裂中期,也是诱导细胞周期同步化的重要方法之一。真核细胞周期可分为4个时期,分别是G1期、S期、G2期和M期。在细胞周期调控中主要有3个控制点,***个控制点在G1期,决定细胞能否进入S期;第二个控制点在G2期,决定细胞能否进入有丝分裂期;第三个控制点在M期,决定细胞是否已经准备好将复制好的染色体拉向两极。CDK(周期蛋白依赖性蛋白激酶)对细胞周期运行起着**性调控作用,CDK与不同时期的周期蛋白结合会在特定周期起调节作用。cyclinA、cyclinB是在M期起调节功能的两种主要周期蛋白。细胞周期运转到分裂中期后,在后期促进复合物(APC)的作用下,M期cyclinA和cyclinB通过泛素化途径迅速降解,Cdkl活性丧失,细胞周期便从M期中期向后期转化。APC活性变化是细胞周期由分裂中期向后期转换的关键因素,其活性受到多种因素的综合调节,纺锤体组装检查点是其重要的调控因素。纺锤体组装不完全,或所有动粒不能被动粒微管全部捕捉,则APC不能被***。 纺锤体微管的动态变化是细胞对外界刺激响应的一部分。深圳无需染色纺锤体胚胎发育
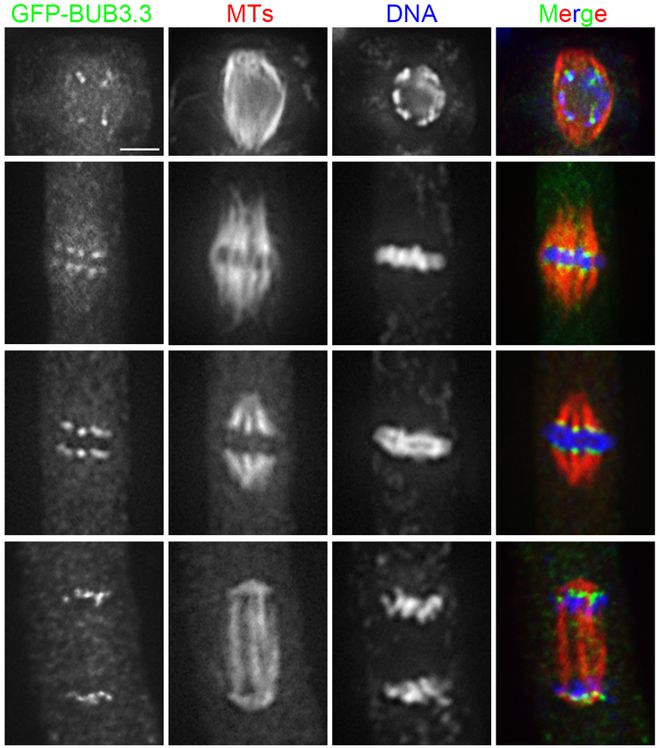
在核移植过程中,纺锤体的稳定性是首要考虑的问题。冷冻和解冻过程中的温度变化和冷冻保护剂的毒性都可能对纺锤体造成损伤,导致染色体分离异常,进而影响胚胎发育。因此,如何在冷冻过程中保持纺锤体的稳定性,是核移植纺锤体卵冷冻研究面临的重要挑战。体细胞核在移入去核卵母细胞后,需要经历复杂的重新编程过程,以获得全能性。然而,这一过程受到多种因素的调控,包括表观遗传修饰、转录因子表达等。在冷冻过程中,这些调控机制可能受到干扰,导致重新编程失败或异常,从而影响胚胎发育。上海成熟卵母细胞纺锤体Hoechst染料纺锤体微管网络的复杂性保证了染色体分离的准确性。
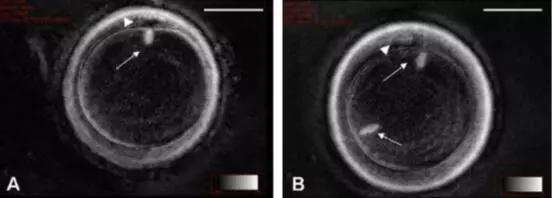
尽管成熟卵母细胞纺锤体冷冻保存技术取得了进展,但仍面临一些挑战。首先,冷冻损伤仍然是制约其临床应用的主要问题之一。尽管玻璃化冷冻法能够在一定程度上减少冷冻损伤,但仍无法完全避免。其次,冷冻保存后的卵母细胞在体外受精和胚胎发育过程中的表现仍存在不确定性。这可能与冷冻过程中纺锤体和染色体的损伤有关,也可能与冷冻保护剂的残留毒性有关。此外,法律伦理问题也是卵母细胞冷冻保存技术面临的一大挑战。不同国家和地区对卵母细胞冷冻保存的法律和伦理规定各不相同,这在一定程度上限制了该技术的普及和应用。
近年来,随着玻璃化冷冻技术的不断发展,成熟卵母细胞纺锤体的冷冻保存研究取得了进展。研究表明,采用玻璃化冷冻法冷冻保存的成熟卵母细胞,在解冻后其纺锤体和染色体的形态及功能均能得到较好的保持。这主要得益于玻璃化冷冻过程中避免了冰晶形成对细胞的损伤,以及冷冻保护剂对细胞的有效保护。然而,值得注意的是,尽管玻璃化冷冻法在提高解冻存活率和妊娠成功率方面取得了成效,但仍存在一些问题。例如,冷冻过程中纺锤体的微管结构可能受到低温的影响而发生解聚,导致染色体分离异常。此外,冷冻保护剂的毒性也可能对卵母细胞造成一定的损伤。为了克服这些问题,研究者们进行了大量的实验和优化工作。例如,通过改进冷冻保护剂的配方和浓度,降低其对细胞的毒性;通过优化冷冻速率和程序,减少冷冻过程中对细胞的机械损伤;以及通过筛选和评估不同冷冻载体和保存时间对卵母细胞冷冻效果的影响,寻找好的冷冻保存条件。纺锤体由微管组成,其动态变化调控着细胞分裂的进程。
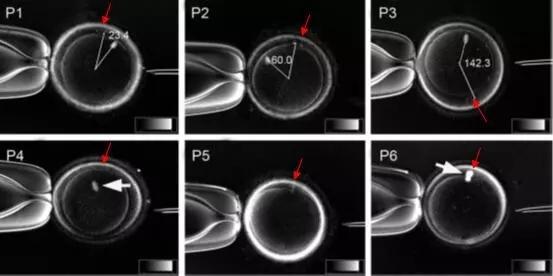
在生殖医学领域,卵母细胞的冷冻保存技术一直是研究的热点,旨在提高女性生育能力的保存与利用。然而,传统的纺锤体观察方法往往需要对卵母细胞进行固定和染色处理,这不*破坏了细胞的活性,还限制了对其发育潜能的深入评估。偏光成像技术,特别是Polscope偏振光显微成像系统,通过利用纺锤体微管结构的双折射性,实现了对纺锤体的无损观察。这种技术无需对卵母细胞进行固定和染色,能够在保持细胞活性的同时,实时、动态地观察纺锤体的形态和变化。这不*提高了观察的准确性和可靠性,还避免了传统染色方法可能带来的细胞损伤和误差。纺锤体在细胞分裂过程中经历明显的形态和结构变化。武汉克隆纺锤体卵细胞评价
纺锤体在细胞分裂中的精确调控是生物体发育的基础。深圳无需染色纺锤体胚胎发育
染色体
当细胞从间期进入有丝分裂期,间期细胞微管网络解聚为游离的αβ-微管蛋白二聚体,再重组成纺锤体,介导染色体的运动;分裂末期纺锤体微管解聚,又重组形成细胞质微管网络。
可分为:动粒微管:连接染色体动粒于两极的微管。
极间微管:从两极发出,在纺锤体中部赤道区相互交错的微管。
星体微管:中心体周围呈辐射分布的微管。
染色体的运动依赖纺锤体微管的组装和去组装。在这一过程中动粒微管与动粒之间的滑动主要是依靠结合在动粒部位的驱动蛋白和动力蛋白沿微管的运动来完成。极微管在纺锤体中部交错,有些分布在极微管之间特殊的双极马达蛋白,其中2个马达蛋白沿一条微管运动,另2个马达结构域沿另一条微管运动。由于2条微管分别来自二极,故极性相反。当双极驱动蛋白四聚体沿微管向正极运动时,纺锤体二极间距离延长。反之纺锤体距离缩短。 深圳无需染色纺锤体胚胎发育